,
The plankton balance hypothesis two opposing forces in the plankton soup nurture and kill By Dr J Floor Anthoni (2003)
www.seafriends.org.nz/enviro/plankton/balance.htm
Plankton
does not only kill by producing poisonous organisms, but it can also kill
because it contains a strong catabolic (decomposing) part consisting of
active bacteria, fungi and viruses that are necessary to decompose dead
organisms and wastes. As a result, most organisms in the sea, and particularly
those who depend on plankton for food, live in constant defiance of death.
By increasing the density of the plankton soup, the additional food is
of no extra value to them, but the decomposing bacteria become a life-threatening
risk. This simple theory (hypothesis) explains many vexing questions about
the undersea ecology and habitat degradation, an ecofactor that has been
overlooked completely.
The mere formulation of a problem is
far more essential than its solution, which may be merely a matter of mathematical
or experimental skills. To raise new questions, new possibilities, to regard
old problems from a new angle requires creative imagination and marks real
advances in science. - Albert Einstein
An introduction to a proposed hypothesis by Floor Anthoni, explaining
why 'innocent' dense plankton blooms can kill a wide range of organisms
over vast areas. June 2003.
When a gradient of plankton density exists along our coasts, an increase
in overall density can cause massive deaths while apparently changing little
to the coastal habitats. However, successive changes are capable of causing
permanent loss, particularly of long-lived species.
Because the juveniles of nearly all species spend some time in the
plankton, feeding from it as they grow, they also expose themselves to
the decomposing component of the plankton. For each exists a most favourable
concentration.
Sunlight is the most important ingredient of life for plants. Marine
plants receive less of it than land plants, so that the availability of
light is more important. Being scarce already, it can be diminished to
disastrously low levels by concentrations of innocent plankton.
Organisms in the sea which have proved to survive attacks by disease-causing
bacteria and viruses may contain potions of benefit to human health and
longevity.
One of the observable differences between underwater life and that
on land, is that major changes in community structure occur over short
distances. A change in rock structure, an overhang, a passage, a niche
or a cave, bring about major changes to what lives there. Why?
Support for this hypothesis came unexpectedly from measurements done
after the 1991-93 kelpbed die-off. It showed not only the extent of the
kelp death but also its severity. Other results will be discussed here
as they arise.
Summing it all up, what the new theory explains and predicts.
.
'
For comments, suggestions and improvements, e-mail
the author, Floor Anthoni.
-- seafriends home -- plankton
index -- Rev 20030626,20030714,20030914,20090406,
Introduction In my search to find answers to the question why we are losing so
much so fast, I have seen the disastrous effects from poisonous plankton
blooms, resulting in mass mortalities of vertebrate species (fish, penguins).
But at the same time in the same localities, the invertebrate species appeared
unaffected. At other times, apparently innocent plankton blooms resulted
in massive loss of invertebrate species, with little effect on vertebrates.
How could this be explained?
Another vexing question asks why marine life is so different at remote
islands compared to the best places along our coast, and why these in turn
differ from locations near population centres. The common theme is that
the water becomes clearer as one travels away from population centres towards
remote islands. But how can this explain their differences in marine life?
Obviously, harmful pollution diminishes but so does nutritious plankton
density.
Then the idea arose that the solution to these and other vexing questions,
is indeed quite simple. Normal plankton feeds and kills at the same time!
It has to, for sound ecological reasons.
The diagram
on right pictures the planktonic food chain, starting from phytoplankton
through zooplankton to higher vertebrates (fish) and mammals like dolphins,
not shown in the diagram. The many trophic (food) levels are necessary
to convert the energy from sunlight into ever larger food packages, from
minuscule plant plankton to zoo plankton to fish larvae to small fish to
large predatory fish. But this is not the complete story. The plankton
food pyramid is part of a larger food chain, a planktonic ecosystem in
which nutrients are eventually restored to feed the phytoplankton. It is
a closed cycle.
This diagram
simplifies the food pyramid above but adds the missing components that
complete the nutrient cycle as is characteristic of any ecosystem. From
nutrients and sunlight the plant plankton captures solar energy and packs
it into carbohydrates and proteins, the basic food for animals, which begin
in the animal plankton and end with higher vertebrates (fish). In the process,
energy is used up for moving and growing, resulting in an ever smaller
biomass (ovals) for each higher trophic level.
As energy is burnt, it is returned in the form of carbondioxide and
water but as animals digest their food, a large part is excreted as waste.
The tiny dead plant plankton and waste particles from zooplankton are intercepted
by the guild of decomposers, consisting of viruses, bacteria and the equivalent
of fungi (saprobic microbes).
Saprophyte: Gk sapros= putrid; phyton=
plant. Sapro- = rotten. Any microorganism living in and on putrid matter
Saprogenic: causing or produced by putrefaction
Saprophile: a bacterium inhabiting putrid matter.
Saprophage: feeding on decaying matter
Decomposition connects
all trophic levels in an ecosystem - Floor Anthoni
Dead phyto- and zooplankton organisms are mostly decomposed in mid water
but the wastes and bodies of larger animals sink to the sea bottom where
they are decomposed by a different guild of benthic (bottom) decomposers.
Some of these have claws and teeth to physically divide the larger particles
into smaller ones. But eventually the nutrients, the ashes of life,
are returned to the water where they take part once again in the planktonic
food cycle.
Because plant plankton has a short life, much of it dies and decomposes
in mid water. As can be seen from the diagram, the amount of energy (food,
dead bodies and waste) flowing through the planktonic decomposers is a
large part of the total. It is a very active guild, and not surprisingly
a threat to life because they decompose living cells as eagerly as dead
ones. Fortunately, the biomass of these decomposers is relatively small
because they are so active, but their numbers are extremely high because
they are so small (picoplankton).
Think for a moment about ourselves. Inside our guts we host a very active
community of decomposers, most of which are bacteria of the genus Eschericia
coli (E coli), a sausage-formed single-celled microbe. We live
in a cautious balance with this destructive organism, protected by a special
slimy lining inside our guts. But things can go wrong as in an ulcer, and
an appendicitis. When the gut bacteria break through the gut lining, entering
the cavity of our belly, death is almost certain as these bacteria decompose
cells they were never meant to decompose. Scratch your skin, and most likely
an infection results from the same guild of bacteria. When left untreated,
such infections can kill.
Now imagine living inside a soup of food and faeces, complete with gut
bacteria, viruses and fungi. That describes what it means to live in the
sea. All marine organisms have developed some level of defence, like our
gut lining, but when overwhelmed by decomposing bacteria and viruses, the
nutritious soup can kill. Thus ironically, plankton has two sides to it:
on the one hand the life-bringing food but on the other hand the deadly
decomposers. It both nurtures and kills. It is thus understandable that
all sea animals have slimy skins to protect themselves from the deadly
decomposers.
The next part of our thesis is the assumption that marine organisms
have evolved to occupy a niche in the gradient from dense to dilute plankton.
If the plankton soup becomes thinner, the organism dies from lack of food.
Conversely, if the soup becomes too thick, organisms can no longer benefit
from the additional food, and they become threatened by the decomposers
instead. So each organism and community of organisms has evolved to live
in balance within a particular density of plankton, most suitable to its
own success. In this manner an unusual plankton bloom can kill a large
area, including many species, without actually being poisonous.
Some organisms have evolved to live in a wide range of conditions and
these we call hardy. Conversely, those restricted to a narrow range are
sensitive. It just so happens that there are many sensitive species but
only few hardy ones.
f033805: In this very degraded environment live three hardy
sponges that are also found in the cleanest of waters: The yellow boring
sponge (Cliona celata), the orange or golden golfball sponge (Tethya
aurantium) and the massive grey sponge (Ancorina alata). (Martins
Bay)
f036831: A fragile and sensitive bryozoan (polyzoan) of the
Pterocella
genus grows on sheltered vertical walls or under overhangs, where currents
flow but waves do not destroy. It is often preyed upon by specialist nudibranchs.
Normally these are clumped together forming a fluffy turf, which makes
photographing an individual difficult. But this photo was taken where nearly
all bryozoa had disappeared, and with them their predators/grazers. This
surviving specimen stands alone and is larger than normal. (Arid Island)
Reader please note that this hypothesis lacks formal proof.
It is attractive as a working hypothesis because it is derived from sound
ecological principles, while it explains many vexing observations. As a
theory it should also be able to predict. To my knowledge, this is the
first time this hypothesis has been proposed. Floor Anthoni, June 2003.
In January 2005 we invented a measuring technique
to quantify the biomass of decomposers and their aggressiveness. Since
then very solid support has been found for the Plankton Balance hypothesis.
See the extensive chapter about the Dark
Decay Analysis (DDA).
This hypothesis was presented at the annual conference
of the New Zealand Marine Sciences Society's in Auckland on 2 Sept 2003
where it was received with hostility and ridicule. The words of session
chair Sam McClatchie, a fisheries scientist from NIWA, sums it all up:
"Although it was an enthusiastic presentation, you have underestimated
your audience and would do well to go back to school to do a course on
plankton ecology".
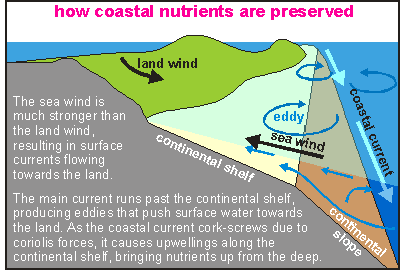
Shifting gradients
The coastal plankton ecosystem can exist only from the nutrients washed
down from the land, from both erosion and eluviation (washing nutrients
out of the soil) and to a lesser extent from the wastes of land animals
(like human sewage). It is an entirely natural process. However, these
days the nutrient density has exceeded what can be called normal by many
times (5 to 30 times). In some places ocean currents add to the coastal
nutrient budget by lifting nutrients from the deep sea to the surface (upwelling).
A plankton
or nutrient gradient results as on the one hand plankton is consumed, while
on the other hand plankton and fine mud ooze down the continental slope
into the abyss (depthlessness) of the the deep ocean. This diagram shows
how soil arrives into the sea as mud. Consisting of three main components,
sands, silt and clay, the mud is winnowed into these components which settle
out in this order away from the shore. Also nutrients are released, giving
rise to plankton blooms a few days later. The plankton cycles renourish
the nutrients, but eventually they are lost down the continental slope.
It is a slow and gradual process acted upon by forces that mop the nutrients
back to the land, thus minimising losses (see diagram)
As a result a gradient exist from thick soup to thin, extending out
from the shore. Similar plankton density gradients exist along the shore,
extending out from dense populations or where rivers enter the sea. Particularly
rivers on large continents provide steep gradients extending outward and
along the shore away from their mouths.
This
diagram shows four gradients. The top two relate to a coastal gradient,
for example when moving away from a population centre along the coast
from say, west to east. The top situation shows where six species A to
F choose to live, at a plankton density most suitable for their existence.
Think of these hypothetical species as hypothetical communities of species.
This is of course a most simplified situation, since many species tolerate
a wider range of plankton densities, such that their zones overlap. As
the coast pollutes, the gradient moves further east. This destroys all
species along the coast, as they become replaced by ones lower in the alphabet.
But it seems as if nothing much has happened but a shift in habitat.
Note how species F becomes extinct as also a new niche is vacated by species
A for unknown (?) species to occupy.
The third and fourth row show a coastal profile, extending outward from
the shore. Species E lives on an island. As the environment pollutes, species
B and E are pushed off their substrate and vanish. Their places become
occupied by species A and D. Again, it seems as if nothing much has happened,
yet major mortalities over the entire region accompanied the process. Species
E may even be pushed to extinction this way if it has no other place to
re-establish itself.
The question marks raise an interesting question: what will establish
in the places left behind by species A? This is where foreign introduced
species may find a foothold. Note that with each increase in pollution,
another habitat shift occurs accompanied by more mass mortalities, and
the possibility for alien species to establish themselves.
In other words, a seemingly innocent increase in pollution will be accompanied
by:
disease, particularly viral, fungal and common infections.
major mortalities over large areas, affecting many species
a chance for alien species to establish themselves, particularly near large
populations of people (!)
loss of long-lived species, particularly sensitive ones
the possibility of true extinctions
This is precisely what we have been observing.
Now imagine that we have a ten year cycle of good years alternated by
bad ones. In other words, the plankton density shifting in and out, east
to west and back again, repeatedly. It will lead to:
permanent loss of many species, including vertebrates (fish)
permanent loss of long-lived species
an overall degradation of all habitats with fewer species, even those far
out in sea over the continental shelf
repeated opportunity for alien species to establish themselves here, because
few if any indigenous species exist to fill this niche.
Again, this is what we have been observing. The consequences of shifting
gradients are indeed rather frightening.
Now imagine the picture with a gradient that is even steeper than shown.
Obviously, the effects will be more accentuated. This is what has been
happening in the past twenty years - reason for alarm.
Gradient in time The above picture is valid not only for spatial gradients, but also
for gradients in time. Wasn't it Capt d'Urville who mentioned in his ship's
log that the water was so clear while anchored where now the Harbour Bridge
in Auckland spans? He could see his anchor 15m down. Visibility there is
now 0.5-1.5m!
Apparently a steep gradient has occurred since the arrival of White
Men, and this gradient has been worsening (steepening) rapidly in the past
twenty years. As shown in the diagram above, this has been accompanied
by mass mortalities, particularly of those species that are attached to
the substrate, unable to move to more suitable places.
This new theory predicts that if we were able to turn back time by cleaning
the water and reducing the concentrations of mud and nutrients, the recovery
phase will also be accompanied by mass mortalities, although less severe.
Reader please note that none of the above has been proved
by scientific method. However, our observations neatly fit the theory.
Killing the babies One of our vexing questions is: why are we seeing so many recruitment
failures? Why are the juveniles so sensitive to degradation? This is an
important question because the juveniles of our commercially caught species,
also spend some time in the plankton as vulnerable larvae.
Most
marine species multiply by releasing eggs and sperm in the water where
they mix and fertilisation takes place. It is a very inefficient way of
reproduction, uncommon with terrestrial species. However in the sea the
overkill in eggs and larvae is necessary to convert the energy from sunlight
into ever larger packages of food. By contrast, land plants are tens of
thousands to millions of times larger than the sea plants that make
up the phyto plankton. On land there is always a plant large enough to
feed the largest organisms (see photo). It can be said that marine organisms
reproduce mainly to make food (99.99%) rather than offspring (0.01%). In
an indirect way, a commercial fish like snapper (Pagrus auratus),
lives from its own spawn.
It is common for marine organisms to start life at less than 1mm in
size, growing to 50mm before leaving their planktonic stage. This equates
to a growth of 50x50x50= 125,000 times, a miraculous act. During that time,
the organism must efficiently find larger food parcels, thus changing its
diet regularly as it grows. How this is achieved for the many species involved,
is not known.
For
our hypothesis it is sufficient to know that nearly all marine organisms,
and certainly all of our commercial species (except sharks, etc) start
life as very small fry (1mm) with proportionally thin skins, being totally
dependent on the composition of the plankton soup. Like other species,
they too are successful only if they are hatched in the right density of
soup with enough nutrition but insufficient decomposition to kill. This
can easily be inadequate, resulting in recruitment failure. For snapper,
temperature is important too. Not because snapper produce more eggs in
warm water, but because for some unknown reasons, warmer water is accompanied
with better hatching and survival conditions for their larvae. Here in
NZ the warm water years are also accompanied by cleaner water.
What we have been seeing underwater is the recruitment failure of many
invertebrate species, and recently also of vertebrate species like snapper.
Scientific fishing trawls confirm this. What is so worrying about this
new problem, is that it is rapidly worsening as we enter a new era of scarcity.
We cannot afford to wait and see.
The shade that kills All plants on Earth depend on sunlight to live. When there is sufficient
light, plants combine carbondioxide and water into carbohydrates, the energy
store for animal life, but also for themselves when sunlight becomes insufficient.
During the night, plants consume the carbohydrates they produced during
the day. Only if their energy budget is positive, will plants grow. Otherwise
they will die. Thus shade can be a profound killer of plants and autotrophic
(self-feeding) microbes.
This
diagram shows the amount of light falling on Earth, for three places on
the northern hemisphere (summer in June/July). The diagram does not take
into account the sunlight intercepted by clouds. Note how light varies
only little from season to season in the tropics (red curve). By comparison
the light at the poles (blue) reaches zero in winter, to rise ABOVE tropical
values in summer, because in summer the sun shines all day. The availability
of light can be a problem for places with dark winters, as the diagram
shows. In a temperate climate like that of New Zealand (about 40º
latitude, green), the difference in incident light between summer (light
intensity x duration= green curve) and winter can be as much as three times
(40/13).
When
light absorption by clouds is taken into account, the difference between
winter and summer becomes larger still. This diagram shows the actual amount
of light measured in Lincoln, New Zealand, which is situated near the middle.
As one can see, the radiation runs from 4 to 24 MJ/m2/day or six times
more light in mid summer than in mid winter. [Guess where the 'winter blues'
come from?]
But for underwater plants the situation becomes worse still due to
the amount of light reflected back into space by the water's surface.
This
diagram shows a polar diagram of the theoretical amount of light reflected
off the water (orange lobe) and that transmitted into the water (blue lobe).
Note that when the sun stands lower than 40 degrees during the day, much
of its light does not enter the sea (one third of blue lobe). For New Zealand
the sun moves in the range of the numbers 4, 5 and 6 in winter and in the
range from 2 to 6 in summer. Note at this point how the light is bent in
such a way that the sun appears to be overhead as the sun rays are descending
more steeply into the water. The blue lobe misses the light from angles
where the words 'transmitted light' appear.
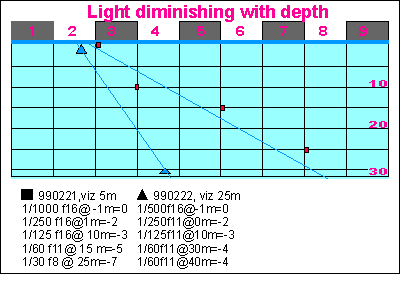
Furthermore, divers observe that the moment their cameras go under
water, they experience a loss of light of at least 50%, which is caused
by the vagaries of ripples and waves. See graph As a result, the light underwater in temperate seas is scarce, a limiting
factor. Not surprisingly, marine algae have evolved methods to catch what
is possible of this scarce amount of light. They have evolved different
pigments like brown and red in order to absorb light more efficiently,
which brings us to the quality of light under water.
This
diagram breaks sunlight down into its visible components and the invisible
UltraViolet A and B. Horizontally it shows wave length (colour) and vertically
the amount absorbed per metre of depth. For ease of reference, coloured
bars show what these wave lengths mean. Three curves are shown, the absorption
in blue oceanic water (50m viz), polluted oceanic water (35m viz) and coastal
water (10m viz). It omits showing the absorption curve for truly polluted
coastal water which lies anywhere above the green curve.
An important conclusion from this graph is that even the clearest of
waters absorb the red and orange components almost as much as coastal water
does. This is simply a property of water. However, note how the UV component
is absorbed quickly by green coastal water compared to clean oceanic water.
But what seaweeds need for growth, are the yellow, green and blue components
of the light. For these the brown and red 'chlorophyll' pigments are most
suitable (chlorophyll is green).
f033429: Most of the large seaweeds (macroalgae) in New Zealand
have brown pigments, like this stalked kelp (Ecklonia radiata) found
here in the sheltered shallows of a rock pool. The brown pigment which
can look reddish, brownish or greenish is suitable for absorbing light
in shallow to moderate depths. (Tawharanui)
f034114: Red seaweeds like this (Pterocladia lucida?)
are usually small. They do well in lower light conditions and deeper water.
Their red pigment is more suitable for absorbing the blue component of
the light, which is more prevalent in the deep. Red seaweeds are sensitive
to pollution. It is not known why. (Mayor I)
f001319: One of the most amazing of marine algae is the
pink paint, a crustose coralline alga (Lithothamnion sp.) or stone-leaf.
It grows a single 'leaf' of hard limestone inside which it lives. This
limestone is hardy enough to survive the bite marks of grazers like sea
urchins, Cooks turban snails and others, all visible in this photo. The
pink paint lives higher on the shore than any other and much deeper too.
(Goat Island)
f034100: Turfing coralline algae have evolved into finely
carved forms like the one shown here, consisting of flexibly jointed bits
of limestone, infused with pink coloured plant life. These plants are very
hardy, resisting strong wave action, low and bright light conditions and
many forms of pollution as well. (Mayor I)
Loss
of light alters coastal marine habitats quite considerably. Whereas seaweeds
are probably less sensitive to the composition of plankton with regards
to its catabolic component (they do not feed on plankton, nor do they breathe
it), they are very sensitive to the loss of light caused by thicker plankton
soup or mud particles in the water.
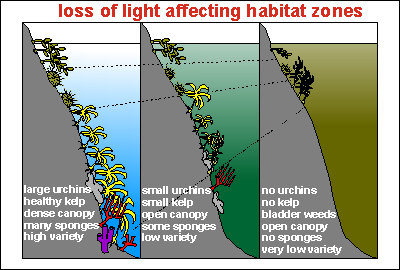
This diagram shows how the coastal fringe degrades. On left a clear
water situation with healthy kelp, dense canopy, large urchins, high biodiversity
and many species of sessile filterfeeders. The photic zone (light zone
suitable for plants) penetrates deep. As the water degrades, so does the
coastal habitat, eventually resulting in very few plant and animal species.
As the photic zone moves up, it is accompanied by a severe loss in biodiversity.
The catabolic component takes care of killing the sessile animal species.
Suffocation from sticky mud is also a problem. The situation with the plankton
soup, however, is quite different.
Plankton species are essentially weightless, living suspended in the
water (this is not entirely true). Eddies and currents move them around
not only horizontally but also vertically (up and down). Whereas a coastal
seaweed lives attached to the rock in a certain position relative to the
photic zone, phytoplankton has the added difficulty of moving in and out
of the photic zone. In other words, the phytoplankton has all the difficulties
of seasonal loss of light plus the problem of being moved in and out of
the photic zone also when the sun shines. Not surprisingly, phytoplankton
suffers high mortality but this is offset by its ability to grow and reproduce
fast, for which sea water temperature can be decisive. What then is the
effect of degrading water quality?
As
the plankton soup becomes thicker or when polluted by clay particles, the
photic zone extends less deep, resulting in poorer growth combined with
higher mortality of the phytoplankton. It is a double-edged sword by which
the productivity of the plankton can diminish very rapidly, leaving the
water free for other organisms like cyanobacteria (bacteria that photosynthesise
like plants) to take over. The soup becomes poisonous. Precisely how this
works is not known, but our theory predicts that the soup becomes less
nutritious and more murderous, resulting in all kinds of unforeseen problems.
Ultimately a dead zone is formed, as happens every year near
the mouth of the Mississippi River in the USA. This diagram shows how sediment
and dense plankton reduce the photic (light) zone. As the overproduction
of phyto plankton rains down to the deep, it dies while decomposers in
the water do their job. More rains down onto the sea soil, where the bottom
decomposers act. They all need oxygen which runs out. Then the anoxic decomposers
take over and the whole sea and bottom become a stinking black putrid mess.
Does the plankton like the rocky shore become less diverse as the plankton
becomes more dense? It behaves like a double-edged sword: on the one side
the threat of reducing light and on the other that of increasing chance
of infection.
Can the plankton become 'sick' such that it no longer functions as
a food producer? Is it possible that the decomposers start to attack the
live producers? It all depends how the balance changes. In the end, the
dead
zones prove that plankton can become very sick indeed. One could even
say that it can die.
Potions for longevity Don't take this chapter very seriously since it is based on speculation,
but our theory predicts that those sessile organisms capable of surviving
the murderous component of the plankton, may indeed contain potions of
benefit to human health and longevity.
Sponges belong to the oldest life forms ever evolved and yet today they
still deserve a place in almost all marine communities. Not being able
to run away or hide, sponges have developed chemical defences from bad
taste to being ultra poisonous. That is why they have attracted interest
from the medical fraternity.
If our theory holds true, then the death and disappearance of sponges
is caused mainly by the murderous component of the plankton, which is not
unlike that of disease-causing microbes and viruses. Being able to survive
their attacks suggests that such sponges have a secret that could well
be of benefit to people. It is an idea worth investigating. Here are the
winners in our contest (also look at the first photo of this section).
f035922: The ugly grey massive sponge (Ancorina alata)
is perhaps the most rugged of all, not only surviving in polluted waters
but also in waters with little nutrition. Here it shows signs of distress,
unable to shrug off the attack by the invading pink paint. (Little Barrier
I)
f011302: These velvety black soft sponges and orange/cream
intestinal seasquirts are typical of highly degraded habitat. Ugly and
ominous, they may well contain potions of longevity. (Houhora Harbour)
Avoidance behaviour Marine organisms can be expected to have evolved avoidance behaviour
to enhance their chances of survival. Those that cannot move around freely,
may choose to close up rather than feed from dense plankton. Such has been
observed in estuarine clams like cockle (Austrovenus [Chione] stutchburyi)
and pipi (Paphies australis) and in the greenlipped mussel (Perna
canaliculus).
Those that are able to roam more freely may opt to limit their feeding
presence in dense plankton, to return to clearer waters for resting and
socialising. Several species choose to spend most of their time in the
clear and cleaner depths below 30m, only to come near the surface for feeding.
Others choose to rest along steep cliff faces where their wastes cannot
collect.
It would be interesting to pursue this idea further to see if some of
the behaviour of marine organisms can be explained in this manner.
Healthy plankton All ecosystems on Earth have evolved over time to make best use of
their resources in protein and energy. They have evolved such that they
convert as much sunlight as they can, while losing as little of their nutrients
as is possible. Such minimum loss or apex ecosystems are usually very stable,
productive and of high biodiversity. Does this also apply to the
plankton ecosystems of which we know so little, and would it be possible
to measure whether a plankton ecosystem is healthy (minimal loss) or sick
(high loss)?
Our new hypothesis is based on the two components of the plankton, the
productive (anabolic) branch and the reductive (catabolic) one. For maximal
productivity, all green matter (phytoplankton of sufficient size) must
be eaten (grazed) by zooplankton, which in turn must be fully predated
on by larger organisms. Only in this way will the amount of waste descending
to the sea bottom, be minimal and also the reductive (disease bringing)
organisms. This is what we would call optimal health.
By contrast, a sick plankton ecosystem would see much of its productivity
in phyto- and zooplankton go to waste. It would have a high reductive component,
bringing a higher risk of disease.
The idea is that the ratio between the two components could be measured
objectively in any sample of seawater. It would consist of sieving the
net plankton (large phytoplankton + small zooplankton) out, and centrifuging
the remainder. The dry biomass of the two compartments could be obtained,
and a ratio established. Likewise the concentrations of oxygen and carbondioxide
could be measured, the one being produced by the producers, the other by
the decomposers.
Assuming that phytoplankton has a short life cycle, one could say by
rule of thumb that a green sea is not being grazed sufficiently, giving
rise to a high disease-bringing component. Visibly green or brown seas
are likely to be also increasing the risk of disease. Thus the simple measure
of visibility, well known to divers, may well be a good indicator of health.
Reader, please note that a simple
and cheap technique (DDA) has been invented to measure the health of
plankton and this can now be done by a 12-year young school student. With
this techniqe we have shown conclusively that plankton can indeed become
sick and murderous and that this kind of plankton has little food value.
Living around the corner A small change in rock form can bring about a large change in the composition
of the invertebrate communities living there. Why? We must distinguish
the different causes: change in sunlight, wave exposure, sediment deposition
and plankton balance.
1) Where the rock face faces the sun light, plants are found because
they compete much better for space than invertebrate organisms. The polar
diagram above shows in the blue lobe that the light underwater comes from
above, even when the sun sets at the horizon. As a result, a steep slope
suddenly loses much of the available light. The difference between sunlit
and shaded sides of rocks is also rather large. It causes large shifts
in plant and animal communities.
2) Wave exposure decreases rapidly with depth but also behind a rock
which shelters its side turned away from the waves. Since large waves always
arrive from the sea, the landward sides of rocks and islands are always
predictably sheltered, allowing more fragile organisms to live.
3) Wave exposure is not always destructive, but may bring salvation
where the environment is threatened by mud. Fine soil particles need calm
water in order to be able to settle out, and wave action prevents this.
Thus the rocks facing moderate wave action are in a better state than those
with complete shelter. Wave action not only prevents sedimentation but
also cleans sediment away.
Because of the above factors, the space underneath overhangs is the
favourite place for sensitive invertebrate life, so easily killed by sedimentation.
But even here the composition of communities can change suddenly.
4) To explain the finer points in what lives where, the plankton balance
hypothesis becomes necessary. In clear waters the availability of food
is important, and invertebrate life favours a place in the current but
sheltered from wave action. The deep reef (below the photic zone) is rich
in invertebrate life where currents prevail, but threatened by sediment
in other places.
Areas of shelter and darkness, lacking currents (caves) can sustain
only the most thrifty of invertebrate life, able to 'live off the smell
of an oily rag'. They do not have adequate reserves to fight disease, so
they are sensitive to the disease effect of denser plankton. In recent
times, these places have also been attacked by unnatural sediment loads,
resulting in direct suffocation and an increase in disease carrying microbes.
As a result, caves and niches in the rock face, once carpeted in life,
now stand barren as a testimony to ongoing degradation. But before sediment
can be seen accumulating in crevices and on ledges, sensitive sponges and
others disappeared because the plankton balance between food and disease
became unfavourable for them.
f036734: This photo shows a sharp boundary between healthy
and sick sponges, due to wave action. Currents rush in the top part of
the photo, keeping the large grey sponge (Anchorina alata) healthy.
But lower down, three of the same species of sponge are dying, For them
the plankton threat was not matched by enough food for fitness to fight
disease. (Arid Island)
f036015: An organ sponge (Callyspongia latituba) stands
dead on a vertical wall, surrounded by other healthy invertebrate species
like orange carpet sponges (Crella incrustans), white anemones (Actinothoe
albocincta) and common sea urchins (Evechinus chloroticus).
This photo was taken during an unusual period of clear water in a year
that saw underwater visibility starkly reduced (May 2003). As a result
of presumably 'sick' plankton conditions, these purple organ sponges were
observed dead and dying. It could be explained that for them the balance
between the good and the bad components in the plankton had tipped unfavourably,
resulting in (slow) death. (Mimiwhangata)
Reader please note that sediment alone is often not a cause of death, but
the bacteria enclosed in it, together with a food source from detritus
and other decomposing materials.
Support for the hypothesis Once
in 1991 and again in 1992 dense plankton blooms reduced the photic zone
so severely and for such a long time that large areas of kelpbed died.
This map shows their extent and severity. During a private expedition measurements
were taken at the sample sites indicated. For more detail see enviro/habitat/survey93.htm.
The 1992/93 kelp dieoff was more severe than that of a year before,
and we observed many organisms dying, apparently because of it. What makes
this event so special is that it happened outside the Hauraki Gulf in an
area of the sea which normally has clear water. The outlying islands and
the north-eastern portion of Great Barrier are not normally threatened
by excessive sedimentation. So the decline in sessile filterfeeders could
be attributed to this single event consisting of a denser than normal plankton
bloom. Amazingly, the plankton bloom was able to kill both plants and animals
in two entirely different ways.
Support
for the Plankton Balance Hypotheses was obtained only recently from our
data of August 1993. Horizontally the severity of the plankton bloom as
measured objectively by the amount of kelp death. To the left none, and
to the right severe. The disappearance of the filterfeeders was measured
by how many were left, in qualitative terms like few and many.
Yet such inaccurate values were adequate for revealing relationships (dose-mortality)
in these cluster diagrams. All sessile filterfeeders were affected by the
plankton bloom, but each at a different rate. Note that the black points
are from places that are influenced by mud and currents, around the Colville
Channel whereas the blue points were subject to a similar plankton
bloom the year before (for details see the survey report).
Actinothoe (white anemone), Polymastia (yellow and orange
nipple sponge), Tethya (pink and golden golfball sponges), Bryozoa
(various species) were all missing where the kelp death was severe. Note
that the pink golfball sponge (Tethya ingalli) proved to be more
sensitive than the orange one (Tethya aurantium).
The plankton balance hypothesis explains why these organisms were affected
and why each has a different sensitivity to such threats.
Eutrophication Processes in Coastal
Systems: Origin and Succession of Plankton Blooms and Effects on Secondary
Production by Robert J. Livingston; Lewis Publishers, Inc.; (November 27, 2000)
Book Description Derived from an unprecedented research effort covering over 31 years
in a series of studies of 7 major river-estuaries, Eutrophication Processes
in Coastal Systems presents a comprehensive and current review of the
nature of the eutrophication process and how short- and long-term nutrient
loading affects marine systems.
This unique book is the culmination of the most advanced research to date
on how coastal systems work. Based on an 11 year interdisciplinary study
of the Perdido Bay System, Dr. Robert J. Livingston's groundbreaking work
offers evidence for significant findings such as:
Nutrient concentration gradients in fresh water as it entered the bay were
stimulatory to phytoplankton blooms
Species that showed distinctive seasonal and interannual successions dominated
plankton blooms
High relative dominance of bloom species was associated with significant
reduction of phytoplankton species richness and diversity
The blooms were associated with major reductions of infaunal and epibenthic
macroinvertibrates, forcing a serious disruption of the food webs and losses
of secondary production
Eutrophication Processes in Coastal Ecosystems goes beyond its innovative
analyses of how estuarine and coastal systems have responded to fundamental
alterations of the eutrophication process. Dr. Livingston's book presents
the case that bloom impacts must be reviewed against the background conditions
that include periodic changes brought on by drought and anthropogenous
dredging. It points to the critical need for further study of phytoplankton
communities and the connection between plankton blooms, sediment deterioration,
and low secondary production.
More indications of support for the hypothesis may follow.
Conclusions The plankton balance hypothesis introduces a new and previously overlooked
limiting factor (ecofactor), the catabolic action of the plankton. Its
main points are:
every drop of seawater is a complete ecosystem: turn the light on and it
makes life; turn it off and it breaks life
plankton both feeds and kills
decomposing microbes can and will cause disease
every marine organism is affected by the density of the plankton
every marine organism lives in a delicate balance between availability
of food and threat of infection
as plankton grows more dense, biodiversity decreases: pelagic, benthic,
planktonic
shifting gradients have serious effects
larvae and juveniles are most affected
as plankton becomes denser, its shading effect makes matters worse
The hypothesis is attractive because it is:
a simple theory/ hypothesis
holistic: precise details are not required
based on sound ecological principles
provides another limiting factor to consider
explains many vexing questions
makes interesting predictions
encounters no conflicts with existing knowledge
Philosophically, the plankton balance hypothesis also has to answer
the fine points why such an important factor has been overlooked by so
many professionals in the field. We made the bold conclusion that a missing
but important limiting factor had to exist to explain the many remaining
paradoxes surrounding species distribution and the ill effects of plankton
density. Then we began to look for one. For this hypothesis to be overlooked,
it had to be:
unique to the aquatic environment - otherwise we would have known about
it.
invisible - otherwise it would have been seen before.
everywhere - because the unexplained paradoxes reigned everywhere
important - otherwise the unexplained paradoxes would be trivial.
simple and obvious - otherwise it would not have been overlooked.
Below follows a point by point summary of the observations explained
by this new theory, and the predictions it makes. Although at the moment
no conflicting observations or facts have been brought to light, they will
in due time also be included as paradoxes contradicting the the hypothesis.
ecosystem gradients and gradients of invertebrate communities
explains biodiversity gradient away from large river mouths
explains biodiversity gradient away from large populations
explains biodiversity gradient away from the shore
explains why remote islands are so different from the the main land.
explains why changes in community structure can happen suddenly (living
around the corner) and in time
erosion and loss of soil; sewage and nutrients from farming; overnourishment
explains why erosion is a major component of degradation
predicts major problems from rapidly increasing erosion in the very recent
past and future
predicts major problems as human habitation densities increase
invertebrate mortalities
explains massive mortalities of many invertebrate species
explains spring mortality, when plankton blooms most densely
explains and predicts cyclical mortalities due to El Niño/ La Niña
or Interdecadal Pacific Oscillation (IPO)
predicts mussel/oyster/salmon farm disease when decomposition on the sea
bottom increases
predicts mussel farm disease when plankton becomes 'sick'
explains that plants are much less affected (this is not quite true, because
some plants are indeed very sensitive)
invasive species, where and when
explains the prevalence of invasive species in the recent past
predicts that this trend is increasing
explains/predicts that invasive alien species establish in the most polluted
places or in places of repetitive mortality
explains/predicts prevalence for a certain phase in the IPO (El Niño
phase)
explains/predicts vertebrate mortalities
vertebrate mortality due to viral, fungal and common infections
explains fish mass mortalities from viral diseases
explains parore (Girella tricuspidata) fungal warts and mortality
predicts an increase in mammal and bird mortalities due to 'normal' infectious
diseases
predicts that animal pathogens are normal plankton catabolic organisms
(viruses, bacteria, fungi)
recruitment failures
predicts recruitment failure due to dense plankton blooms
predicts recruitment failure in certain phases of the IPO cycle (El Niño)
explains mussel spat recruitment problems
predicts that recruitment failures will become more common and more serious
aquariums/ biology
explains the kinds of disease found in aquariums
explains why so many marine creatures have slimy skins
explains why damage to the skin's slime layer can cause infections
visible indicators
explains that underwater visibility (viz) is a major indicator
explains that sediment (mud/clay) increases the catabolic component
explains that sturdy and sensitive organisms are both good indicator species
potions for longevity
predicts that chemicals beneficial for health are present in hardy organisms,
particularly those surviving extreme degradation and those living inside
caves
paradoxes seemingly contradicting the plankton balance hypothesis
 Plankton
does not only kill by producing poisonous organisms, but it can also kill
because it contains a strong catabolic (decomposing) part consisting of
active bacteria, fungi and viruses that are necessary to decompose dead
organisms and wastes. As a result, most organisms in the sea, and particularly
those who depend on plankton for food, live in constant defiance of death.
By increasing the density of the plankton soup, the additional food is
of no extra value to them, but the decomposing bacteria become a life-threatening
risk. This simple theory (hypothesis) explains many vexing questions about
the undersea ecology and habitat degradation, an ecofactor that has been
overlooked completely.
Plankton
does not only kill by producing poisonous organisms, but it can also kill
because it contains a strong catabolic (decomposing) part consisting of
active bacteria, fungi and viruses that are necessary to decompose dead
organisms and wastes. As a result, most organisms in the sea, and particularly
those who depend on plankton for food, live in constant defiance of death.
By increasing the density of the plankton soup, the additional food is
of no extra value to them, but the decomposing bacteria become a life-threatening
risk. This simple theory (hypothesis) explains many vexing questions about
the undersea ecology and habitat degradation, an ecofactor that has been
overlooked completely.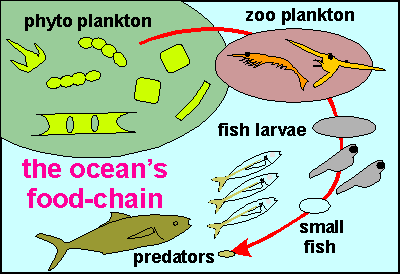 The diagram
on right pictures the planktonic food chain, starting from phytoplankton
through zooplankton to higher vertebrates (fish) and mammals like dolphins,
not shown in the diagram. The many trophic (food) levels are necessary
to convert the energy from sunlight into ever larger food packages, from
minuscule plant plankton to zoo plankton to fish larvae to small fish to
large predatory fish. But this is not the complete story. The plankton
food pyramid is part of a larger food chain, a planktonic ecosystem in
which nutrients are eventually restored to feed the phytoplankton. It is
a closed cycle.
The diagram
on right pictures the planktonic food chain, starting from phytoplankton
through zooplankton to higher vertebrates (fish) and mammals like dolphins,
not shown in the diagram. The many trophic (food) levels are necessary
to convert the energy from sunlight into ever larger food packages, from
minuscule plant plankton to zoo plankton to fish larvae to small fish to
large predatory fish. But this is not the complete story. The plankton
food pyramid is part of a larger food chain, a planktonic ecosystem in
which nutrients are eventually restored to feed the phytoplankton. It is
a closed cycle.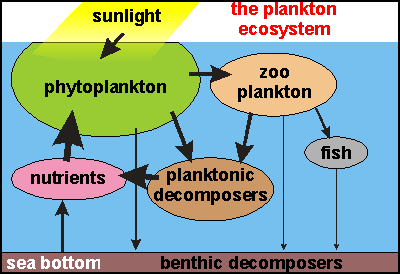 This diagram
simplifies the food pyramid above but adds the missing components that
complete the nutrient cycle as is characteristic of any ecosystem. From
nutrients and sunlight the plant plankton captures solar energy and packs
it into carbohydrates and proteins, the basic food for animals, which begin
in the animal plankton and end with higher vertebrates (fish). In the process,
energy is used up for moving and growing, resulting in an ever smaller
biomass (ovals) for each higher trophic level.
This diagram
simplifies the food pyramid above but adds the missing components that
complete the nutrient cycle as is characteristic of any ecosystem. From
nutrients and sunlight the plant plankton captures solar energy and packs
it into carbohydrates and proteins, the basic food for animals, which begin
in the animal plankton and end with higher vertebrates (fish). In the process,
energy is used up for moving and growing, resulting in an ever smaller
biomass (ovals) for each higher trophic level.


 A plankton
or nutrient gradient results as on the one hand plankton is consumed, while
on the other hand plankton and fine mud ooze down the continental slope
into the abyss (depthlessness) of the the deep ocean. This diagram shows
how soil arrives into the sea as mud. Consisting of three main components,
sands, silt and clay, the mud is winnowed into these components which settle
out in this order away from the shore. Also nutrients are released, giving
rise to plankton blooms a few days later. The plankton cycles renourish
the nutrients, but eventually they are lost down the continental slope.
It is a slow and gradual process acted upon by forces that mop the nutrients
back to the land, thus minimising losses (see
A plankton
or nutrient gradient results as on the one hand plankton is consumed, while
on the other hand plankton and fine mud ooze down the continental slope
into the abyss (depthlessness) of the the deep ocean. This diagram shows
how soil arrives into the sea as mud. Consisting of three main components,
sands, silt and clay, the mud is winnowed into these components which settle
out in this order away from the shore. Also nutrients are released, giving
rise to plankton blooms a few days later. The plankton cycles renourish
the nutrients, but eventually they are lost down the continental slope.
It is a slow and gradual process acted upon by forces that mop the nutrients
back to the land, thus minimising losses (see 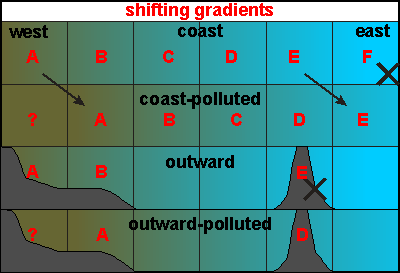 This
diagram shows four gradients. The top two relate to a coastal gradient,
for example when moving away from a population centre along the coast
from say, west to east. The top situation shows where six species A to
F choose to live, at a plankton density most suitable for their existence.
Think of these hypothetical species as hypothetical communities of species.
This is of course a most simplified situation, since many species tolerate
a wider range of plankton densities, such that their zones overlap. As
the coast pollutes, the gradient moves further east. This destroys all
species along the coast, as they become replaced by ones lower in the alphabet.
But it seems as if nothing much has happened but a shift in habitat.
Note how species F becomes extinct as also a new niche is vacated by species
A for unknown (?) species to occupy.
This
diagram shows four gradients. The top two relate to a coastal gradient,
for example when moving away from a population centre along the coast
from say, west to east. The top situation shows where six species A to
F choose to live, at a plankton density most suitable for their existence.
Think of these hypothetical species as hypothetical communities of species.
This is of course a most simplified situation, since many species tolerate
a wider range of plankton densities, such that their zones overlap. As
the coast pollutes, the gradient moves further east. This destroys all
species along the coast, as they become replaced by ones lower in the alphabet.
But it seems as if nothing much has happened but a shift in habitat.
Note how species F becomes extinct as also a new niche is vacated by species
A for unknown (?) species to occupy. Most
marine species multiply by releasing eggs and sperm in the water where
they mix and fertilisation takes place. It is a very inefficient way of
reproduction, uncommon with terrestrial species. However in the sea the
overkill in eggs and larvae is necessary to convert the energy from sunlight
into ever larger packages of food. By contrast, land plants are tens of
thousands to millions of times larger than the sea plants that make
up the phyto plankton. On land there is always a plant large enough to
feed the largest organisms (see photo). It can be said that marine organisms
reproduce mainly to make food (99.99%) rather than offspring (0.01%). In
an indirect way, a commercial fish like snapper (Pagrus auratus),
lives from its own spawn.
Most
marine species multiply by releasing eggs and sperm in the water where
they mix and fertilisation takes place. It is a very inefficient way of
reproduction, uncommon with terrestrial species. However in the sea the
overkill in eggs and larvae is necessary to convert the energy from sunlight
into ever larger packages of food. By contrast, land plants are tens of
thousands to millions of times larger than the sea plants that make
up the phyto plankton. On land there is always a plant large enough to
feed the largest organisms (see photo). It can be said that marine organisms
reproduce mainly to make food (99.99%) rather than offspring (0.01%). In
an indirect way, a commercial fish like snapper (Pagrus auratus),
lives from its own spawn.
 For
our hypothesis it is sufficient to know that nearly all marine organisms,
and certainly all of our commercial species (except sharks, etc) start
life as very small fry (1mm) with proportionally thin skins, being totally
dependent on the composition of the plankton soup. Like other species,
they too are successful only if they are hatched in the right density of
soup with enough nutrition but insufficient decomposition to kill. This
can easily be inadequate, resulting in recruitment failure. For snapper,
temperature is important too. Not because snapper produce more eggs in
warm water, but because for some unknown reasons, warmer water is accompanied
with better hatching and survival conditions for their larvae. Here in
NZ the warm water years are also accompanied by cleaner water.
For
our hypothesis it is sufficient to know that nearly all marine organisms,
and certainly all of our commercial species (except sharks, etc) start
life as very small fry (1mm) with proportionally thin skins, being totally
dependent on the composition of the plankton soup. Like other species,
they too are successful only if they are hatched in the right density of
soup with enough nutrition but insufficient decomposition to kill. This
can easily be inadequate, resulting in recruitment failure. For snapper,
temperature is important too. Not because snapper produce more eggs in
warm water, but because for some unknown reasons, warmer water is accompanied
with better hatching and survival conditions for their larvae. Here in
NZ the warm water years are also accompanied by cleaner water.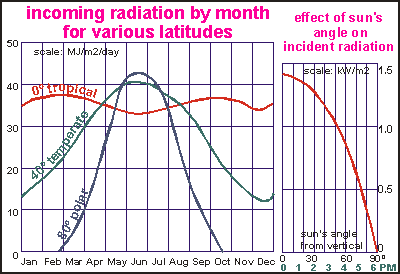 This
diagram shows the amount of light falling on Earth, for three places on
the northern hemisphere (summer in June/July). The diagram does not take
into account the sunlight intercepted by clouds. Note how light varies
only little from season to season in the tropics (red curve). By comparison
the light at the poles (blue) reaches zero in winter, to rise ABOVE tropical
values in summer, because in summer the sun shines all day. The availability
of light can be a problem for places with dark winters, as the diagram
shows. In a temperate climate like that of New Zealand (about 40º
latitude, green), the difference in incident light between summer (light
intensity x duration= green curve) and winter can be as much as three times
(40/13).
This
diagram shows the amount of light falling on Earth, for three places on
the northern hemisphere (summer in June/July). The diagram does not take
into account the sunlight intercepted by clouds. Note how light varies
only little from season to season in the tropics (red curve). By comparison
the light at the poles (blue) reaches zero in winter, to rise ABOVE tropical
values in summer, because in summer the sun shines all day. The availability
of light can be a problem for places with dark winters, as the diagram
shows. In a temperate climate like that of New Zealand (about 40º
latitude, green), the difference in incident light between summer (light
intensity x duration= green curve) and winter can be as much as three times
(40/13).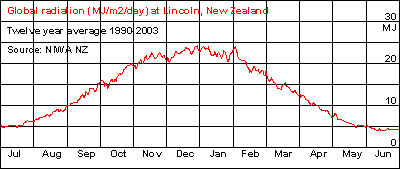 When
light absorption by clouds is taken into account, the difference between
winter and summer becomes larger still. This diagram shows the actual amount
of light measured in Lincoln, New Zealand, which is situated near the middle.
As one can see, the radiation runs from 4 to 24 MJ/m2/day or six times
more light in mid summer than in mid winter. [Guess where the 'winter blues'
come from?]
When
light absorption by clouds is taken into account, the difference between
winter and summer becomes larger still. This diagram shows the actual amount
of light measured in Lincoln, New Zealand, which is situated near the middle.
As one can see, the radiation runs from 4 to 24 MJ/m2/day or six times
more light in mid summer than in mid winter. [Guess where the 'winter blues'
come from?]
 This
diagram shows a polar diagram of the theoretical amount of light reflected
off the water (orange lobe) and that transmitted into the water (blue lobe).
Note that when the sun stands lower than 40 degrees during the day, much
of its light does not enter the sea (one third of blue lobe). For New Zealand
the sun moves in the range of the numbers 4, 5 and 6 in winter and in the
range from 2 to 6 in summer. Note at this point how the light is bent in
such a way that the sun appears to be overhead as the sun rays are descending
more steeply into the water. The blue lobe misses the light from angles
where the words 'transmitted light' appear.
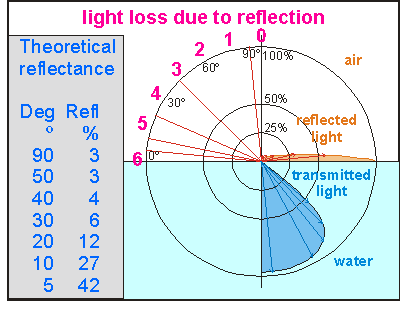
This
diagram shows a polar diagram of the theoretical amount of light reflected
off the water (orange lobe) and that transmitted into the water (blue lobe).
Note that when the sun stands lower than 40 degrees during the day, much
of its light does not enter the sea (one third of blue lobe). For New Zealand
the sun moves in the range of the numbers 4, 5 and 6 in winter and in the
range from 2 to 6 in summer. Note at this point how the light is bent in
such a way that the sun appears to be overhead as the sun rays are descending
more steeply into the water. The blue lobe misses the light from angles
where the words 'transmitted light' appear.
 This
diagram breaks sunlight down into its visible components and the invisible
UltraViolet A and B. Horizontally it shows wave length (colour) and vertically
the amount absorbed per metre of depth. For ease of reference, coloured
bars show what these wave lengths mean. Three curves are shown, the absorption
in blue oceanic water (50m viz), polluted oceanic water (35m viz) and coastal
water (10m viz). It omits showing the absorption curve for truly polluted
coastal water which lies anywhere above the green curve.
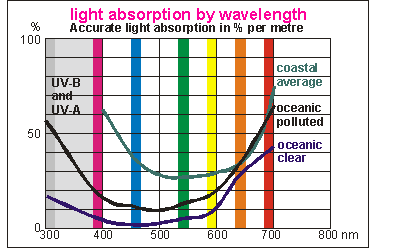
This
diagram breaks sunlight down into its visible components and the invisible
UltraViolet A and B. Horizontally it shows wave length (colour) and vertically
the amount absorbed per metre of depth. For ease of reference, coloured
bars show what these wave lengths mean. Three curves are shown, the absorption
in blue oceanic water (50m viz), polluted oceanic water (35m viz) and coastal
water (10m viz). It omits showing the absorption curve for truly polluted
coastal water which lies anywhere above the green curve.




 Loss
of light alters coastal marine habitats quite considerably. Whereas seaweeds
are probably less sensitive to the composition of plankton with regards
to its catabolic component (they do not feed on plankton, nor do they breathe
it), they are very sensitive to the loss of light caused by thicker plankton
soup or mud particles in the water.
Loss
of light alters coastal marine habitats quite considerably. Whereas seaweeds
are probably less sensitive to the composition of plankton with regards
to its catabolic component (they do not feed on plankton, nor do they breathe
it), they are very sensitive to the loss of light caused by thicker plankton
soup or mud particles in the water.
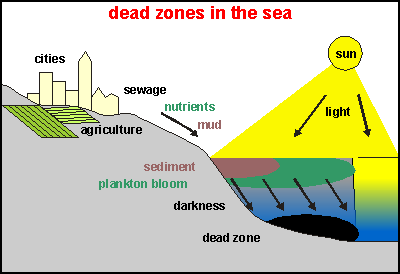 As
the plankton soup becomes thicker or when polluted by clay particles, the
photic zone extends less deep, resulting in poorer growth combined with
higher mortality of the phytoplankton. It is a double-edged sword by which
the productivity of the plankton can diminish very rapidly, leaving the
water free for other organisms like cyanobacteria (bacteria that photosynthesise
like plants) to take over. The soup becomes poisonous. Precisely how this
works is not known, but our theory predicts that the soup becomes less
nutritious and more murderous, resulting in all kinds of unforeseen problems.
As
the plankton soup becomes thicker or when polluted by clay particles, the
photic zone extends less deep, resulting in poorer growth combined with
higher mortality of the phytoplankton. It is a double-edged sword by which
the productivity of the plankton can diminish very rapidly, leaving the
water free for other organisms like cyanobacteria (bacteria that photosynthesise
like plants) to take over. The soup becomes poisonous. Precisely how this
works is not known, but our theory predicts that the soup becomes less
nutritious and more murderous, resulting in all kinds of unforeseen problems.




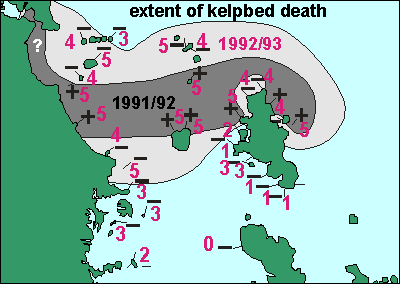 Once
in 1991 and again in 1992 dense plankton blooms reduced the photic zone
so severely and for such a long time that large areas of kelpbed died.
This map shows their extent and severity. During a private expedition measurements
were taken at the sample sites indicated. For more detail see
Once
in 1991 and again in 1992 dense plankton blooms reduced the photic zone
so severely and for such a long time that large areas of kelpbed died.
This map shows their extent and severity. During a private expedition measurements
were taken at the sample sites indicated. For more detail see 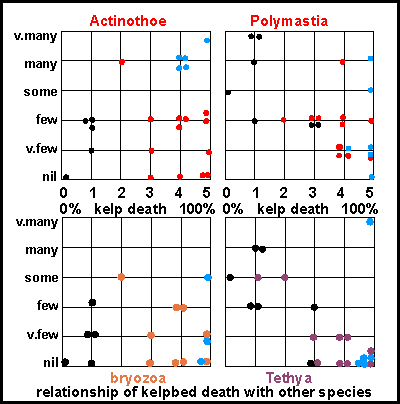 Support
for the Plankton Balance Hypotheses was obtained only recently from our
data of August 1993. Horizontally the severity of the plankton bloom as
measured objectively by the amount of kelp death. To the left none, and
to the right severe. The disappearance of the filterfeeders was measured
by how many were left, in qualitative terms like few and many.
Yet such inaccurate values were adequate for revealing relationships (dose-mortality)
in these cluster diagrams. All sessile filterfeeders were affected by the
plankton bloom, but each at a different rate. Note that the black points
are from places that are influenced by mud and currents, around the Colville
Channel whereas the blue points were subject to a similar plankton
bloom the year before (for details see the survey report).
Support
for the Plankton Balance Hypotheses was obtained only recently from our
data of August 1993. Horizontally the severity of the plankton bloom as
measured objectively by the amount of kelp death. To the left none, and
to the right severe. The disappearance of the filterfeeders was measured
by how many were left, in qualitative terms like few and many.
Yet such inaccurate values were adequate for revealing relationships (dose-mortality)
in these cluster diagrams. All sessile filterfeeders were affected by the
plankton bloom, but each at a different rate. Note that the black points
are from places that are influenced by mud and currents, around the Colville
Channel whereas the blue points were subject to a similar plankton
bloom the year before (for details see the survey report).
{kind=link}
{kind=link}